Colour Lesson X
Afterimages: the eye that outlasts the scene
Every decided colour does a certain violence to the eye, and forces the organ to opposition.
Lesson IX showed that a colour’s neighbours change its appearance. The surround pushes the centre toward its complement. This lesson asks what happens when the neighbour is removed entirely. The eye, having been pressed in one direction for long enough, does not simply return to neutral. It overcorrects. It sees the complement where there is nothing: a ghost of the colour that just left.
The ghost in the lamplight
Goethe was sitting at an inn when a girl with a brilliantly coloured complexion entered the room. He looked at her in the bright lamplight, then glanced away at the opposite white wall. There, on the blank plaster, he saw a dark face surrounded by a glowing light. The figure had the same form, but her warm tones had become their opposites.
He had encountered a negative afterimage : a visual impression persisting after the stimulus has gone, in the complementary colour of the original.
This was not an isolated curiosity. Goethe spent years cataloguing such phenomena. He noticed that staring at red left a cyan-green ghost. Staring at yellow left a violet ghost. Staring at blue left an orange ghost. The pattern was consistent.
The complement is not an abstraction invented by the colour wheel. It is the physiological residue of the original colour, left behind in the eye.Red
Cyan afterimage
Yellow
Violet afterimage
Blue
Orange afterimage
Goethe’s interpretation was not quite right. He believed the eye was seeking a kind of wholeness, completing what was missing. But his observations were precise, and the pattern he recorded pointed directly toward the opponent structure of colour vision, which would take another century to describe properly.
What cone fatigue actually is
The retina contains three classes of cone photoreceptors: long-wavelength (L), medium-wavelength (M), and short-wavelength (S). Each is sensitive to a different region of the spectrum, with peak sensitivities roughly corresponding to red-orange , green , and blue-violet .
When you stare at a red surface for thirty seconds or more, the L-cones in that region of the retina are stimulated heavily and continuously. The photopigments in those cones bleach. The cones become desensitised, temporarily less responsive to long-wavelength light.
When you then look at a neutral white wall, the wall reflects all wavelengths roughly equally. But the L-cones in that retinal region are fatigued. They fire less. The M and S cones, which had been relatively unstimulated, fire at full strength. The result: the signals reaching the brain are shifted away from red and toward the cone combination that normally signals cyan-green . You see the complement, not because it is there, but because the machinery for detecting the original colour has worn itself out.
The afterimage is not a vision of what is there. It is a vision of what the eye can no longer suppress.The effect fades as the photopigments regenerate, typically over ten to thirty seconds. It is stronger when the stimulus was more saturated, more prolonged, and more narrowly localised on the retina.
Hering’s opponent channels
Young and Helmholtz established the trichromatic theory: three cone types encode the initial signal. But trichromacy alone cannot fully explain afterimages. Why does fatiguing the red response produce exactly cyan? Why not a random drift?
The answer came from Ewald Hering in 1892. Hering proposed that the visual system processes colour not in three independent channels but in three opponent pairs : red against green , yellow against blue , and white against black .
Each pair is a see-saw: stimulating one end suppresses the other. Red and green cannot be perceived simultaneously in the same place at the same intensity. Neither can yellow and blue. This is why there is no such thing as reddish-green or yellowish-blue. The opponent pairs are mutually exclusive at the neural level.
There are colours whose contraries are completely opposed to each other... They mutually destroy each other, and produce, when mixed, a neutral gray or white.
When you fatigue the red end of the red-green channel, the green end fires unopposed. That is the afterimage colour. The see-saw tips.
Both theories are correct at different stages of the visual pathway. Trichromacy operates at the photoreceptors. Opponent processing begins at the ganglion cells in the retina and continues through the lateral geniculate nucleus into the visual cortex. The two accounts are not competing. They describe consecutive stages of the same system.
Young-Helmholtz is the hardware. Hering is the firmware. The afterimage is where firmware takes over.Positive afterimages
Not all afterimages are negative. A very brief, very bright stimulus can produce a positive afterimage : a lingering sensation in the same colour as the original, not its complement.
Look directly at a camera flash for a fraction of a second. For several seconds afterward, you see the shape of the flash, positive, in roughly the same colour. This is not cone fatigue but the continuation of the original neural discharge: the photoreceptors briefly maintain their activation even after the stimulus ends.
Positive afterimages are short-lived, lasting a second or two at most, and they give way to the negative complement as cone fatigue sets in. The progression: same colour briefly, then complement as the fatigue dominates.
This persistence of neural activity is the mechanism behind persistence of vision : the illusion that a rapid sequence of still images is continuous motion. A film frame is replaced by darkness between exposures, but the positive afterimage of the previous frame bridges the gap. Cinema is built on the brief stubbornness of the visual system.
The positive afterimage holds the frame. The negative afterimage pays the debt afterward.The surgeon’s scrubs
The most striking practical application of chromatic afterimages is in the operating theatre.
Surgeons spend hours looking at red and pink tissue . The L-cones in the foveal region of their retinas are chronically stimulated during a long procedure. When they glance away from the surgical field to check a monitor, look at a colleague, or rest their eyes, they see blue-green afterimage ghosts wherever they look. On a white wall or a colleague’s white coat , those ghosts appear vividly.
Operating room walls and surgical scrubs are now almost universally blue-green . The logic is opponent process. A blue-green background provides very low contrast against the blue-green afterimage. Instead of a vivid ghost appearing on a neutral field, the afterimage merges into the environment. The visual noise is reduced. The surgeon can look away without losing the ability to see clearly when looking back.
The colour of a surgeon's scrubs is not a uniform code or a brand choice. It is opponent process theory applied to error reduction in high-stakes work.The same logic governs the green-tinted glass used in some operating room lights, the desaturated wall colours in radiology reading rooms, and the recommendation that photo editors work in neutral grey environments. The surround is managed so that the eye arrives at its next look without a pre-loaded bias.
Afterimages in art
Artists have understood this effect, even without the neuroscience to name it.
Georges Seurat’s pointillism places small dots of pure colour adjacent to each other, letting the eye blend them optically. At the right viewing distance, brief chromatic afterimages from one dot influence the apparent colour of its neighbours. The technique exploits both simultaneous contrast (Lesson IX) and the residual afterimage between saccades.
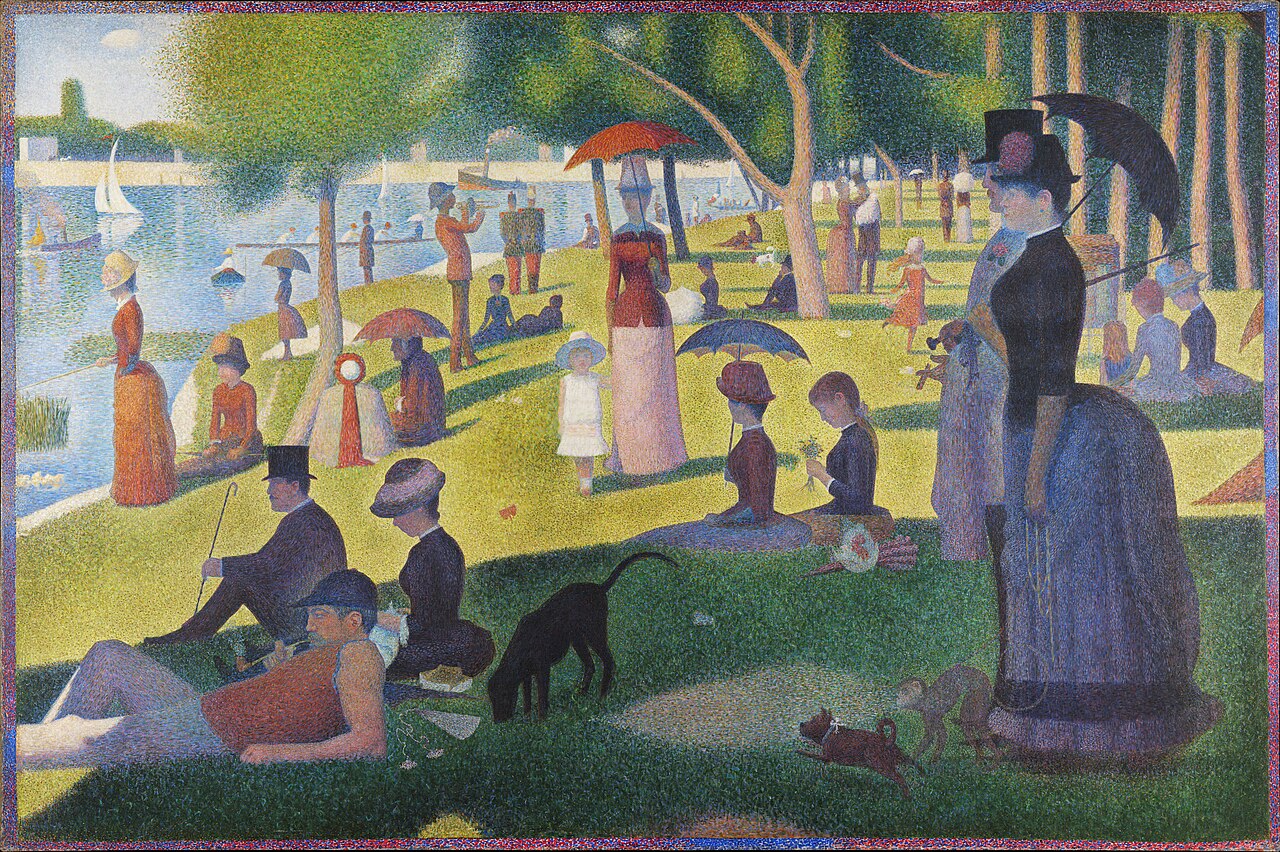
Bridget Riley’s Op Art works in similar territory. Her blue and white stripe paintings from the 1960s produce a shimmering quality partly because the eye, fatigued by a blue stripe, moves to a white stripe and briefly sees a faint orange tint. The flicker is in the eye, not in the pigment.
Riley spoke of wanting the painting to happen in the viewer’s eye. The canvas is a score. The performance is in the retina.
When an artwork vibrates, it is often the eye vibrating in response. The artist has composed a stimulus that the visual system cannot resolve into stillness.What this means for design work
Designers who work with colour for extended periods are not always seeing what they think they are seeing.
Staring at a saturated palette for an hour shifts your adaptation state. The cones most stimulated by your primary colour fatigue progressively. When you look at the rest of your design, every neutral area acquires a faint cast of the complement. What seems like a neutral grey background may be carrying a subtle bias that disappears when a fresh eye looks at the file.
This is the physiological argument behind the “fresh eyes” rule: step away before a final colour decision. It is not a superstition. It is cone regeneration.
Colour calibration workflows account for this. Display calibration is done after a period of dark adaptation, in a controlled neutral environment, so that the operator’s eye is not pre-loaded with any particular bias. Print proofing rooms are built around neutral grey walls for the same reason.
The designer's eye is not a neutral instrument. It is a fatigued, adapted, history-carrying perceiver. Managing that is part of the craft.The practical implication: when making a colour call that matters, change the environment. Step into a different light. Look at something neutral for thirty seconds. Let the cones reset. Then look again.
Closing thought
Lessons VII through X form a quartet of perceptual effects. Metamerism: two surfaces that match under one light diverge under another. Colour constancy: the brain corrects for illuminant shift to hold surface identity stable. Simultaneous contrast: the neighbour pushes the centre toward its complement. Afterimage: the neighbour leaves, and the eye pushes itself toward the complement anyway.
In each case, the lesson is the same. There is no such thing as a colour read in isolation. Every perception is conditioned by what came before, what is alongside, and what the eye most recently believed to be true.
The eye is not a recorder. It is an interpreter. And like any interpreter, it brings its own history to every reading.After the individual perception effects, the question turns outward: not how one colour affects another by accident, but how colours can be deliberately arranged to agree with each other. That is harmony, and it is where we go next.
References
- Johann Wolfgang von Goethe, Theory of Colours (1810) - Ewald Hering, Outlines of a Theory of the Light Sense (1892) - Rudolf Arnheim, Art and Visual Perception (1954) - John E. Dowling, The Retina: An Approachable Part of the Brain - Josef Albers, Interaction of Color - David Briggs, huevaluechroma.com - Bruce MacEvoy, handprint.com
Write something in the light
Leave a thought, reflection, or a quiet ripple below.